
2007年,来自丹尼斯克公司(一家总部位于丹麦哥本哈根的食品添加剂公司,目前被杜邦公司收购)的科学家找到了一种能增强细菌防御噬菌体能力的方法。之后2013年,四个研究团队报告了这一被称为CRISPR的系统,自此CRISPR技术红红火火的发展了起来,许多科研团队利用它来删除、添加、激活或抑制人体、老鼠、斑马鱼、细菌、果蝇、酵母、线虫和农作物细胞中的目标基因。
近期来自蒙大拿州立大学的两位学者以“CRISPR-RNA-Guided Adaptive Immune Systems”为题,介绍了CRISPR-Cas免疫应答系统。其中Blake Wiedenheft就是当年加州大学伯克利分校Jennifer Doudna研究组成员,他们曾于2010年破解了Csy4核糖核酸内切酶原子水平的晶体结构模型——研究人员确定Csy4是一种存在于原核细胞的酶,它能够启动生成CRISPR衍生RNAs (crRNAs),这种小RNA分子能够靶向并沉默侵入的病毒和质粒。
细菌和古细菌进化出了复杂的CRISPR适应性免疫系统,这一系统可以分成I-III三个类型,至少有11种不同的亚型:从I-A到I-F,从II-A到II-C,以及III-A和III-B,不过尽管有这些不同,所有的CRISPR-Cas系统都通过三个主要阶段来完成功能:采集,CRISPR RNA(crRNA) 生物合成与靶向干扰。
阶段1:外源DNA采集
外源核苷酸是通过Cas蛋白来识别的,入侵的短片段DNA(30-50个碱基对)被称为protospacers,作为间隔序列插入宿主CRISPR位点中,由重复序列隔开。对于I型和II型系统来说,protospacers来自入侵DNA中两侧出现2-5个核酸结构(PAM,protospacer adjacent motif)的区域。一般protospacers连接在CRISPR 位点的一端,并且后者通过涉及Cas1、Cas2和游离3'-hydroxyls等元件的机制,牵引序列。Protospacer的整合过程中也出现了牵引末端重新序列复制,这可能涉及宿主聚合酶和DNA修复机制。
相关论文解析:
Multiple mechanisms for CRISPR–Cas inhibition by anti-CRISPR proteins
研究人员确定了其中三种anti-CRISPR蛋白:AcrF1、AcrF2和AcrF3的功能机制。他们对这些蛋白进行了生物化学及体内研究,证实每一个anti-CRISPR蛋白都通过不同的机制来抑制了CRISPR–Cas的活性。有两个anti-CRISPR阻断了CRISPR–Cas复合物的DNA结合活性,但它们是通过与不同的蛋白质亚基互作,利用了空间或非空间抑制模式来做到这一点的。第三种anti-CRISPR蛋白通过结合Cas3解螺旋酶-核酸酶,阻止其招募到结合DNA的CRISPR–Cas复合物上来起作用。在体内,这一anti-CRISPR可以将CRISPR–Cas系统转变为转录遏制物,首次证实了一种蛋白质相互作用蛋白可以调控CRISPR–Cas活性。作者们认为,这些anti-CRISPR蛋白质不同的序列及作用机制表明了独立的进化,并预示了还存在其他的方式——蛋白质借助于它们改变了CRISPR–Cas的功能。
新研究首次探讨了蛋白质抑制CRISPR–Cas系统的机制。这些多样且不同的机制反映了病毒-宿主军备竞赛深层的进化根源。这些已知和尚有待发现的Anti-CRISPR,将为认识和操控CRISPR–Cas系统提供大量有价值的工具。其中一个例子就是,新研究发现AcrF3通过阻止招募Cas3将CRISPR–Cas系统转变为了一个基因调控因子。由于除了破坏外源DNA,CRISPR–Cas系统来执行着各种功能,许多重要的功能有可能是由与CRISPR–Cas元件互作,由此改变了这一系统活性的蛋白质来完成。
阶段2:crRNA合成CRISPR RNA
生物合成在转录之后,生成初级转录产物:pre-crRNA,之后经过加工,又成为一组短小的CRISPR 衍生RNAs(crRNAs),这些crRNAs每一个都包含有对应于之前遇到的外源DNA的对应序列。
CrRNAs导向序列两端是相邻重复序列区域,在I型和II型系统中,这种CRISPR转录产物会被CRISPR特异性核酸内切酶(Cas6 或 Cas5d)切割,切割位点位于重新序列。许多I型系统的重新序列会出现多次,因此Cas6也需要稳定连接在crRNA 3‘端茎环上。对于III型系统来说,Cas6则是短暂连接,crRNA 3’端会进一步通过未知的酶处理。
II型系统中,CRISPR RNA加工过程则取决于反式作用 crRNA (tracrRNA),tracrRNA包含一个重复序列的互补序列,这些双螺旋区域在Cas9出现时可以通过RNase III 进行处理。
相关论文解析:
CRISPR-mediated adaptive immune systems in bacteria and archaea.
这篇发表在Annu. Rev. Biochem. 杂志上的文章十分重要,解析了crRNA合成过程,以及其后的靶向干扰中的几个重要步骤,此后也被多人引用。
Development and Applications of CRISPR-Cas9 for Genome Engineering张锋的这篇综述概述了CRISPR/Cas9作为一种平台技术的开发状况以及在基因组编辑方面的应用,也讨论了其存在的一些挑战,以及未来的创新之路。
阶段3:靶向干扰
成熟的crRNAs能指导Cas蛋白靶向互补靶标,靶标序列由专用Cas 核酸酶降解,但其靶标降解的机制存在差异。I型和II型系统都可以靶向包含PAM和protospacer互补序列的dsDNA底物。III型系统则不依赖于 PAM作为识别序列,而是通过导向序列延伸至crRNA信号5'handle的核苷酸进行识别(CRISPR位点包含与导向序列,5'handle互补的序列),并阻止靶向切割。
相关论文解析:
Co-transcriptional DNA and RNA Cleavage during Type III CRISPR-Cas Immunity
一些数据表明,当病毒入侵细菌细胞时,称作为III型CRISPR-Cas的这一机制会靶向病毒的DNA,阻止它利用细菌的机器来拷贝自身及感染更多的细菌。但另外的一些实验表明,III型CRISPR-Cas只能通过切割病毒RNA来让病毒丧失能力。
洛克菲勒大学的研究人员他们检测了III型CRISPR-Cas对DNA和RNA的切割,结果发现了从前其他人没有得到过的一个关键成分,并发现CRISPR-Cas确实切割了病毒DNA生成的RNA,但它也切割了病毒的DNA。
这种双交叉系统有一些优势。许多的病毒整合到它们感染细胞的基因组中保持休眠状态,不会造成损伤。事实上,这些病毒对于细菌可能是有益的,例如它们携带的毒素帮助了细菌促进自身生存。举例说来,白喉毒素是由一种细菌所分泌,但编码这一毒素的基因却来自于一种病毒。只有在病毒开始将它们的DNA转录为RNA之时才会让它们丧失功能,通过设置这样的要求III型CRISPR-Cas不会损及休眠病毒,使得它们可以继续让宿主细菌受益。
循环关闭I型系统中,监测复合物中靶向结合会导致Cas3介导的靶向降解(直接干扰)或最初采集,这其中涉及crRNA导向募集Cas3、Cas1 和Cas2到外源DNA处,引起新一轮的快速采集。
II型系统中虽然未观察到最初采集,但Cas9 也是protospacer筛选的必要元件,这表明在靶向干扰与外源DNA采集之间存在一种功能性联系。近期还有研究发现了编码anti-CRISPRs 蛋白的不同病毒基因,这也是指出了干扰以上不同阶段,能颠覆CRISPRs 系统。
Cas9 specifies functional viral targets during CRISPR-Cas adaptation.
一些证据表明,某些Cas酶(未包括Cas9)自身可以操控记忆形成过程。基于Cas9识别切割位点的方式,研究人员猜测Cas9在记忆形成中也发挥了作用。
除了匹配CRISPR引导序列和病毒DNA,Cas9需要在附近寻找第二信号:病毒DNA中的前间区序列邻近基序( protospacer adjacent motif,PAM)序列。这是一个至关重要的步骤,因为PAM序列的存在阻止了Cas9攻击细菌自身包含记忆的DNA。
为了检验他们的假说,研究人员交换了化脓链球菌和嗜热链球菌免疫系统的Cas9酶,它们各自识别不同的PAM序列。结果,PAM序列跟随着在两种细菌之间发生了交换——表明在记忆形成中Cas9负责了PAM的识别。
Multiple mechanisms for CRISPR–Cas inhibition by anti-CRISPR proteins研究人员确定了其中三种anti-CRISPR蛋白:AcrF1、AcrF2和AcrF3的功能机制。他们对这些蛋白进行了生物化学及体内研究,证实每一个anti-CRISPR蛋白都通过不同的机制来抑制了CRISPR–Cas的活性。有两个anti-CRISPR阻断了CRISPR–Cas复合物的DNA结合活性,但它们是通过与不同的蛋白质亚基互作,利用了空间或非空间抑制模式来做到这一点的。第三种anti-CRISPR蛋白通过结合Cas3解螺旋酶-核酸酶,阻止其招募到结合DNA的CRISPR–Cas复合物上来起作用。在体内,这一anti-CRISPR可以将CRISPR–Cas系统转变为转录遏制物,首次证实了一种蛋白质相互作用蛋白可以调控CRISPR–Cas活性。作者们认为,这些anti-CRISPR蛋白质不同的序列及作用机制表明了独立的进化,并预示了还存在其他的方式——蛋白质借助于它们改变了CRISPR–Cas的功能。
新研究首次探讨了蛋白质抑制CRISPR–Cas系统的机制。这些多样且不同的机制反映了病毒-宿主军备竞赛深层的进化根源。这些已知和尚有待发现的Anti-CRISPR,将为认识和操控CRISPR–Cas系统提供大量有价值的工具。其中一个例子就是,新研究发现AcrF3通过阻止招募Cas3将CRISPR–Cas系统转变为了一个基因调控因子。由于除了破坏外源DNA,CRISPR–Cas系统来执行着各种功能,许多重要的功能有可能是由与CRISPR–Cas元件互作,由此改变了这一系统活性的蛋白质来完成。
来源:生物通

为你推荐
 资讯
资讯 国家药监局批准两款创新医疗器械
近日,国家药监局批准两款创新医疗器械上市。分别为苏州无双医疗设备有限公司的植入式心律转复除颤器和深圳大医伽玛刀科技有限公司的头部伽玛射线立体定向放射治疗系统。
2026-07-15 18:34
 资讯
资讯 翰森制药最高23亿美元授权并参与纳斯达克上市公司股权投资
7月14日,翰森制药发布公告宣布,已与Avere Therapeutics, Inc (Avere)签订独家许可协议及参与战略投资。
2026-07-15 14:20
 资讯
资讯 2026年底前至少完成一轮集中采购,国家卫生健康委 国家中医药管理局 国家疾病预防控制局关于进一步做好公立医疗卫生机构医用设备集中采购工作
国家级集中采购。国家卫生健康委继续负责组织全国公立医疗卫生机构甲类大型医用设备集中采购。个性化定制程度高、需配套设计并建设基础设施工程、逐台按程序获得注册的设备,待...
2026-07-15 09:27
 资讯
资讯 中国医药工业主营业务收入100强
近日,由中国医药工业信息中心举办的2026第43届全国医药工业信息年会在北京开幕,备受关注的“2025年度中国医药工业主营业务收入前100位企业”同步揭晓。
2026-07-14 19:55
 资讯
资讯 国家药监局批准1类创新药阿更葡糖钠注射液上市
近日,国家药品监督管理局批准杭州奥默医药股份有限公司申报的1类创新药阿更葡糖钠注射液(商品名称:奥美克松)上市,该药品用于拮抗罗库溴铵诱导的神经肌肉阻滞。该药品上市为...
2026-07-14 17:01
 资讯
资讯 迪哲医药最高15亿美元BD阿斯利康
今日,迪哲医药发布公告,公司授予阿斯利康在全球范围内的独家开发、商业化舒沃哲的权利。公司将获得阿斯利康支付的一次性、不可返还的首付款6亿美元,最高达4亿美元的临床开发...
2026-07-14 14:57
 资讯
资讯 国家医保局、国家药监局开展跨部门联合检查
近日,国家医保局、国家药监局组成联合检查组,依据药品追溯码筛查线索对内蒙古自治区和山西省部分地市医药机构、药品批发企业开展联合检查。国家医保局党组成员、副局长黄华...
2026-07-14 11:46
 资讯
资讯 2026年国家医保目录及商保目录调整通过形式审查名单公布
2026年目录调整申报阶段,国家医保局共收到基本医保目录申报信息800份,涉及药品通用名664个,最终601个通过形式审查,其中目录外368个,目录内233个,总体通过率91%。共收到商...
2026-07-14 10:34


 资讯
资讯 全国首笔药品追溯码无追索权保理业务在温州落地
近日,在温州市医保局的积极推动下,全国首笔基于药品追溯码的无追索权保理业务成功落地。国药控股温州有限公司凭借药品追溯码形成的真实交易记录,将其对温州医科大学附属眼视...
2026-07-13 10:15

 资讯
资讯 时隔8年调整的2026版国家基药目录新增的4种1类创新药和独家品种
7月9日,国家卫生健康委、国家中医药局、国家疾控局三部门公布2026年版国家基本药物目录。国家基本药物目录在新医改启动后,于2009、2012、2018年完成三轮更新,这是时隔8年后的...
2026-07-12 21:31
 资讯
资讯 陈薇院士出任中国工程院党组成员、副院长
据中国工程院官网消息,根据中共中央和国务院通知,中国工程院第九届院领导班子组成如下:张玉卓同志任党组书记、院长;张军、陈杰、李仲平、陈建峰、陈薇等同志任党组成员、副...
2026-07-11 21:36

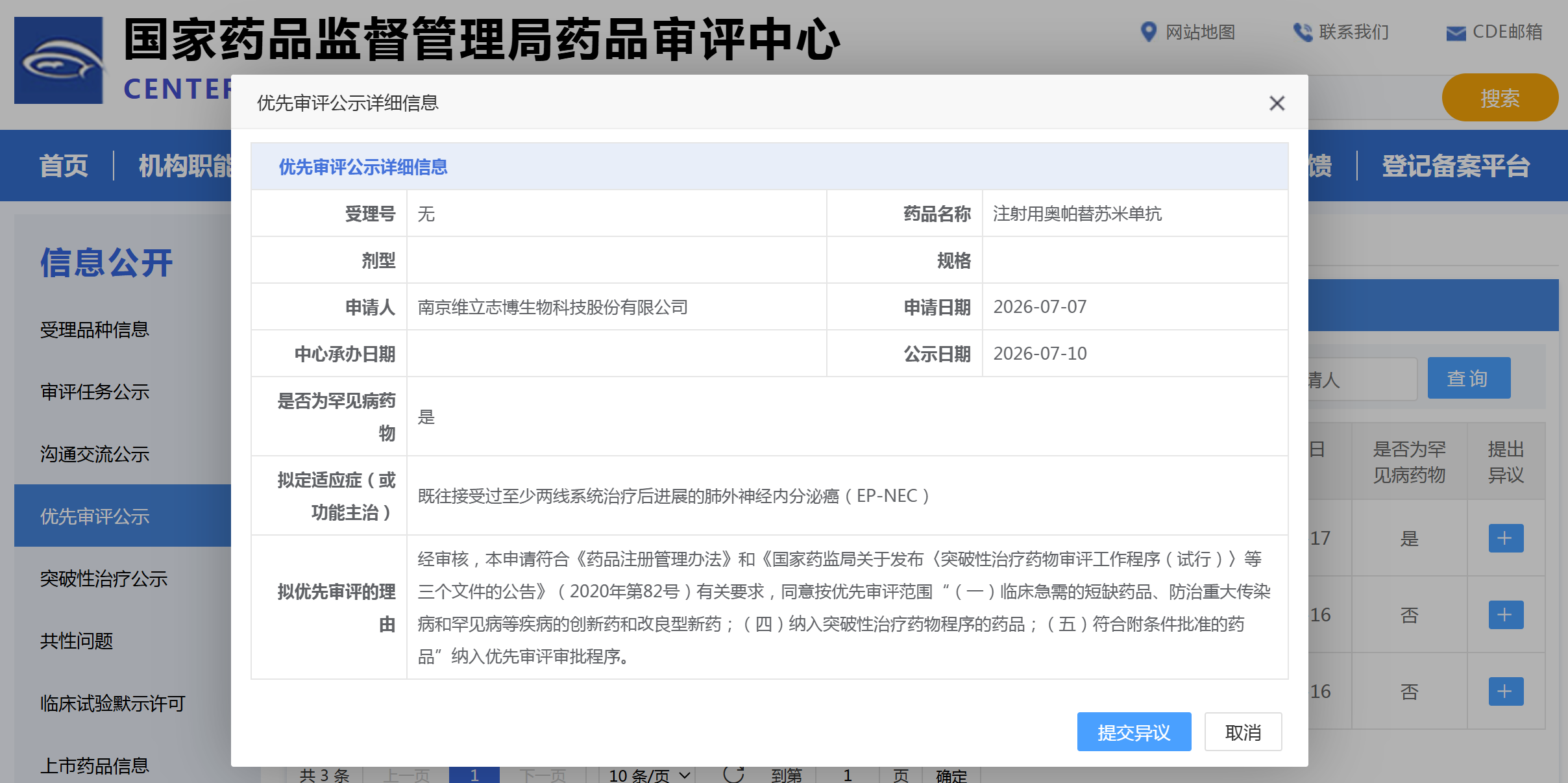 资讯
资讯 维立志博PD-L1/4-1BB 双抗拟纳入优先审评
7月10日,国家药监局药品审评中心官网显示,南京维立志博生物自主研发的注射用奥帕替苏米单抗拟纳入优先审评品种,申报适应症为既往接受过至少两线系统治疗后进展的肺外神经内分...
2026-07-10 15:08
 资讯
资讯 丰原药业实控人拟变更为安徽蚌埠市国资委
7月9日,丰原药业发布公告,公司控股股东安徽丰原集团有限公司的一致行动人7月8日与蚌埠投资集团有限公司签署《股份转让意向协议》,蚌埠投资集团拟受让丰原集团的一致行动人持...
2026-07-10 13:49